
Pinus jeffreyi Grev. & Balf.
Jeffrey Pine
Pinaceae -- Pine family
By: James L. Jenkinson
From: Burns, Russell M., and Barbara H. Honkala, tech. coords. 1990. Silvics of North America: 1. Conifers; Agriculture Handbook 654. U.S. Department of Agriculture, Forest Service, Washington, DC. vol.2, 877 p.
Jeffrey pine (Pinus jeffreyi) was discovered in 1852 in the Shasta Valley of California by John Jeffrey, a Scottish botanical explorer. Partly overlapping ponderosa pine (Pinus ponderosa) in range and superficially resembling it, Jeffrey pine was first classified as a variety of ponderosa pine (28,45). These western yellow pines produce wood of identical structure and quality and are closely related taxonomically (10). Jeffrey pine is distinct chemically, ecologically, and physiologically and is readily distinguished from ponderosa pine on the basis of bark, leader, needle, bud, and cone morphology (23).
Habitat
Native Range
Primarily a California species, Jeffrey pine ranges north through the Klamath Mountains into southwestern Oregon, across the Sierra Nevada into western Nevada, and south in the Transverse and Peninsular Ranges into northern Baja California (10,20). This distribution is intimately linked with edaphic factors in the northwest portion of the range and strongly reflects climatic and elevational factors in the northeast, central, and southern portions.
Jeffrey pine thrives in comparatively harsh environments throughout most of its range (1,21,24,55,58). Contrasted with ponderosa pine, Jeffrey pine completes annual top growth sooner, enters dormancy earlier, and requires longer cold exposure for leader growth in spring (27). Because Jeffrey pine is especially cold hardy (21,22), tolerant of drought (51,58,59), and adapted to short growing seasons, it competes well and typically dominates other conifers on cold, xeric, and infertile sites.
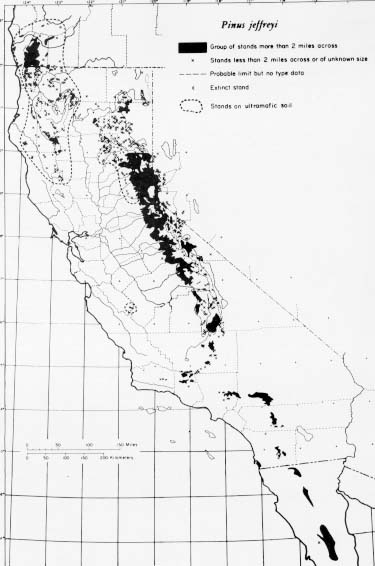
- The native range of Jeffrey pine.
Climate
Jeffrey pine grows well in diverse temperature regimes. Cold winters largely distinguish its range east of the Sierra-Cascade crest from that in the Klamath Mountains, western Sierra Nevada, and southern California. Most populations east of the crest are exposed to January mean minima between -13° and -5° C (8° and 23° F), while those in the west and south are between -7° and 2° C (19° and 36° F).
Summer nights are warmer and differences in day-night temperatures are smaller for Jeffrey pine in the western Sierra Nevada and southern California than elsewhere in the range. July day-night differences in these regions are as little as 11° C (20° F), and rarely exceed 19° C (34° F). In the Klamath Mountains and east of the Sierra-Cascade crest, July day-night differences mostly exceed 19° C (34° F), approach 26° C (47° F) in some areas, and decrease to 13° C (24° F) only for stands at highest elevations.
Throughout the range, precipitation falls mostly during the winter season. Mean annual rainfall averages as little as 380 to 430 mm (15 to 17 in) in places east of the Sierra-Cascade crest, is only 200 mm. (8 in) for certain stands scattered along the eastern Sierra Nevada and Virginia Range of western Nevada (3), and exceeds 1270 to 1520 mm (50 to 60 in) in parts of the western Sierra Nevada and Klamath Mountains. Mean snowfall in Jeffrey pine localities typically ranges from 30 cm (12 in) or less at lowest elevations in the Klamath Mountains to well over 520 cm (204 in) at high elevations in the Sierra Nevada, particularly along the central crest.
Soils and Topography
Perhaps one-fifth of the distribution of Jeffrey pine is on ultramafic soils. At middle elevations on the western slope of the northern Sierra Nevada and in the North Coast Range and Klamath Mountains, Jeffrey pine often dominates and is almost entirely restricted to soils derived from ultramafic rocks- peridotites and their alteration products, serpentinites. The typical forest soils formed on such rocks are fine, fine loamy, and clayey texture skeletal surface soils. On these highly infertile, mostly shallow soils, Jeffrey pine descends to low elevations: 490 m (1,600 ft) in Butte County, 260 m (850 ft) in Humboldt County, 60 m (200 ft) in Del Norte County, CA, and 183 m (600 ft) in Douglas County, OR. The only native Jeffrey pine in California's South Coast Range grows on an isolated mass of sterile serpentine in San Benito County (19). Jeffrey pine's innately short growing season, limited nutrient and water demands, and extensive root growth probably ensure its presence on poor sites.
Above 1600 m (5,250 ft) in ultramafic regions and at all elevations everywhere else within its range, Jeffrey pine grows on any well-drained forest soil, regardless of parent material. Most of the usual soils that carry Jeffrey pine are coarse or gravelly sandy loams or loamy coarse sands that often merge with rocklands. Where these soils are of recent volcanic origin (ashes, pumice, or cinders) they are Dystric or Typic Xerorthents or Xeropsamments of the order Entisols. Jeffrey pine characteristically grows on granitic soils in the Sierra Nevada. These soils extend over complex systems of branching ridges, ravines, and canyons at elevations ranging from 1520 to 2740 m (5,000 to 9,000 ft), are usually found in a humid microthermal climate, and vary from about 0.6 to 1.4 m (2 to 5 ft) deep. They are Dystric and Typic Xerochrepts and Xerumbrepts of the order Inceptisols. Volcanic soils and mud flows commonly support Jeffrey pine in the northern Sierra Nevada and adjacent Cascade Range. They are extensive on gentle to steep slopes of dissected plateau-like areas at elevations up to 1830 m (6,000 ft), and most vary from 0.5 to 1 m (1.6 to 3.3 ft) deep (31). They are Ultic, Typic, and Dystric Haploxeralfs of the order Alfisols.
East of the Sierra Nevada crest from Lassen to Alpine Counties, CA, and along the Virginia Range of adjacent western Nevada, isolated patches of Jeffrey and ponderosa pines grow on more than 125 islands of altered andesite (3). These edaphically restricted stands range in elevation from 1310 to 2130 m (4,300 to 7,000 ft), and many are within the Pinyon-Juniper woodland (20).
Associated Forest Cover
Incense-cedar (Libocedrus decurrens) is the most widespread associate of Jeffrey pine on ultramafic soils. Locally prominent are Douglas-fir (Pseudotsuga menziesii), Port-Orford-cedar (Chamaecyparis lawsoniana), ponderosa pine, sugar pine (Pinus lambertiana), western white pine (P. monticola), knob-cone pine (P. attenuata), Digger pine (P. sabiniana), and Sargent cypress (Cupressus sargentii). Above 1600 m (5,250 ft) in the Klamath Mountains, North Coast Range, and northern Sierra Nevada, Jeffrey pine shares various soils and sites with California red fir (Abies magnifica), white fir (A. concolor), sugar pine, incense-cedar, western white pine, and Sierra lodgepole pine (Pinus contorta var. murrayana).
South of the Pit River in northeastern California and on the east side of the Cascade Range in southwestern Oregon and northern California, Jeffrey and ponderosa pines form extensive forests and usually intermingle in both closed and open, parklike stands. Jeffrey pine forests range widely from 1520 to 2130 m (5,000 to 7,000 ft) of elevation in the northern Sierra Nevada, and from 1830 to 2900 m (6,000 to 9,500 ft) in the central and southern Sierra Nevada. Ponderosa pine, sugar pine, white fir, incense-cedar, California red fir, western white pine, lodgepole pine, and western juniper (Juniperus occidentalis) all mix in locally, but few of them join Jeffrey pine on south slopes and granitic soils (9,44).
Jeffrey pine is the dominant yellow pine in forests east of the Sierra Nevada crest and in the Transverse and Peninsular Ranges into Baja California. In the Sierra San Pedro Martir, it ranges from 1830 to 3050 m (6,000 to 10,000 ft) and shares the southern limits of sugar pine, white fir, incense-cedar, and lodgepole pine (10,45,52).
Jeffrey pine forests constitute one of the more unusual forest cover types in western North America (14). Because Jeffrey pine has wide edaphic and elevational ranges in diverse physiographic regions, Jeffrey Pine (Society of American Foresters Type 247) is highly variable and adjoins or merges with many others: Red Fir (Type 207), White Fir (Type 211), Lodgepole Pine (Type 218), Pacific Douglas-Fir (Type 229), Port-Orford-Cedar (Type 231), Douglas-Fir-Tanoak-Pacific Madrone (Type 234), Interior Ponderosa Pine (Type 237), Western Juniper (Type 238), Pinyon-Juniper (Type 239), Sierra Nevada Mixed Conifer (Type 243), Pacific Ponderosa Pine-Douglas-Fir (Type 244), Knob-cone Pine (Type 248), Canyon Live Oak (Type 249), and California Mixed Subalpine (Type 256). Associated understory species are diverse, reflecting climatic influences (14, p. 123).
Life History
Our present understanding of Jeffrey pine's life history is probably limited, because most studies have concentrated on populations in northern California.
Reproduction and Early Growth
Flowering and Fruiting- In California, Jeffrey pine flowers in June or July, depending on the climatic region, elevation, aspect, and annual variation in temperature (12). The species is monoecious. Female strobili emerge from the bud shortly after the male strobili. After pollination, the conelets develop slowly, reaching less than one-fifth the size of mature cones the first growing season. Fertilization occurs about 13 months after pollination, and the cones grow rapidly to reach full size in summer of the second season.
Unripe cones are pale or dark purple to black and shade to light brown or dull purple at maturity. Seeds are mature when cone specific gravity (fresh weight basis) drops to between 0.81 and 0.86; they are safely collected in stands where one or two trees have cones that are cracking, with seed scales separating. Mature cones are usually 13 to 23 cm (5 to 9 in) long, open to resemble old-fashioned straw beehives or skeps, and normally shed most of their seeds in September or October (30).
Seed Production and Dissemination- Although trees as young as 8 years have borne a cone crop, typical cone-bearing Jeffrey pines are 18 to 55 m (60 to 180 ft) tall and produce a large seed crop every 2 to 8 years (30).
When shed, the winged seeds drop about 2.2 m/s (7.2 ft/s) (49), indicating that winds of 8 km/h (5 mi/h) carry them no further than the height of seedfall. Seeds can be widely spread, however. Fall storms are common in California's mountains, and winds average 13 to 26 km/h (8 to 16 mi/h) in September and October. Gusts occasionally exceed 64 to 113 km/h (40 to 70 mi/h), enough to blow seeds up to 15 times the height of seedfall, even 750 m (2,460 ft) from a tree height of 50 m (164 ft).
Besides wind, certain seed eaters also disseminate seeds. In the Sierra Nevada, Clark's nutcracker harvests and stores substantial quantities of ripe Jeffrey pine seeds, burying them in many small clusters in a wide variety of microsites, and often where snow accumulates least and melts rapidly in spring (54). At least eight other common birds also extract and eat seeds of Jeffrey pine.
Several kinds of squirrels cut and store large quantities of Jeffrey cones for their seeds, including the widespread golden-mantled ground squirrel and western gray squirrel. The chickaree cuts whole cones and buries them in the ground, and chipmunks harvest seeds by gnawing cones in the tree. Mice and voles efficiently gather, cache, and consume large quantities of shed seed.
Like seeds of most pines in temperate climates, Jeffrey pine germinates quickly the spring after seedfall. For starting new stands after harvest, however, natural regeneration is seldom quick and never reliable. The usual cause of failure is vegetation in the original understory that simply preempts the site. Irregular seed crops, poor seed dissemination, seed predators, cutworms, pathogens, mammals, and drought are also lethal factors (16,25).
Seedling Development- Mature seeds show highly variable degrees of dormancy after air drying and cold storage. Different seed sources require different amounts of moist, cold stratification for rapid and complete germination (30). Although certain sources east of the Sierra-Cascade crest may not require pretreatment, stored seeds of most sources germinate best after 60 days of stratification.
In forest tree nurseries in northern California and southern Oregon, seeds are sown in April to utilize the full growing season. Germination is epigeal (30). Stratified seeds are sown above a maximum depth of 6 to 8 mm (0.25 to 0.30 in) and at a density to produce 269 to 323 seedlings per square meter (25 to 30/ft²) . Fertilization and irrigation regimes are tailored to seedling requirements in the particular nursery soil and climate. For most sources west of the Sierra-Cascade crest and in southern California, seedlings of plantable size are raised in one growing season. For many sources east of the crest, seedlings are often carried through two seasons.
To consistently raise large and healthy seedlings of Jeffrey pine, nursery soil management is crucial. In midsummer, dry, fallow soil is ripped deeply enough to restore rapid drainage and aeration, and then fumigated. Fumigation is necessary to control nematodes, root rots such as Rhizoctonia, Phytophthora, Pythium, Macrophomina, and Fusarium spp., and foliar diseases such as Phoma and Sirococcus (38).
Jeffrey pine is quickly established in the field when dormant seedlings are lifted from nursery beds at the right time in winter, held in cold storage, planted at the right time in spring, and protected against animal damage and competing vegetation. Lifted seedlings are root-pruned 23 cm (9 in) below the cotyledon scars and stored in polyethylene-lined bags at 1° C (34° F). Planting starts at the onset of spring conditions, when soils warm sufficiently to permit water uptake and root growth. Planting is ideally completed before the last spring rain to ensure that roots will be sealed in the soil. In the Sierra Nevada, field survivals of 90 to 99 percent are attainable on cleared sites within the species' elevational range (27).
Vegetative Reproduction- Jeffrey pine does not sprout.
Sapling and Pole Stages to Maturity
Growth and Yield- In field plantations, Jeffrey pine grows less rapidly than ponderosa pine during the sapling stage, but more rapidly in the pole stage. Jeffrey pine has trailed ponderosa pine in height growth through more than 5 years in the northern Sierra Nevada in Plumas County, CA (15), 6 years in the Warner Mountains in Modoc County, CA (35), and 11 years in the western Sierra Nevada in Stanislaus County, CA (table 1).
Species |
Plantation Age | Tree Height | Stem diameter | Leader length | Height increase |
| yr | m | cm | m | pct | |
| Jeffrey pine | 11 | 2.15 | 4.3 | 0.44 | 26 |
| 20 | 7.05 | 16.2 | 0.56 | 9 | |
| Ponderosa pine | 11 | 3.64 | 8.9 | 0.43 | 13 |
| 20 | 7.19 | 17.8 | 0.49 | 7 | |
| yr | ft | in | ft | pct | |
| Jeffrey pine | 11 | 7.1 | 1.7 | 1.4 | 26 |
| 20 | 23.1 | 6.4 | 1.8 | 9 | |
| Ponderosa pine | 11 | 11.9 | 3.5 | 1.4 | 13 |
| 20 | 23.6 | 7.0 | 1.6 | 7 | |
¹Seedlings were planted on cleared sites at an elevation of about 1650 m (5,400 ft) in 1962. |
|||||
After 5 years in the Plumas test, every stock class of Jeffrey pine gradually overtook its ponderosa counterpart, averaging 127 cm (50 in) tall and exceeding ponderosa pine by 5 percent 9 years after planting (15). In the Stanislaus plantation, Jeffrey pine accelerated growth into the pole stage, increasing height by one-fourth at 10 years, and at 20 years had nearly overtaken ponderosa pine in both height and diameter (table 1). Leader length at 20 years was 14 percent greater for Jeffrey pine, indicating that the species' difference might soon be eliminated. In the Modoc plantation, trees averaged 5 m (16.5 ft) tall and 15 cm (6 in) in diameter when thinned at 30 years (34,35). At thinning and for the next 15 years, the growth of these poles was apparently the same for both species. When seedlings of Jeffrey and ponderosa pines from the Sierra Nevada were planted at 560 m (1,830 ft) in the North Coast Range in Mendocino County, CA, the Jeffrey pines outgrew ponderosa pines from comparable elevations, edging them in both height and diameter in 17 years (6).
Jeffrey pine may live 400 to 500 years and on the best sites can reach an impressive size. Trees larger than 152 cm (60 in) in d.b.h. were often measured in virgin forests east and west of the Sierra-Cascade crest. The largest known survivor is on the Stanislaus National Forest in the western Sierra Nevada and measures 229 cm (90 in) in diameter and 53 m (175 ft) tall (36). Yellow pines taller than 61 m (200 ft) are recorded in early volume table measurements, and some of them probably are Jeffrey pine (24).
Several general accounts state that Jeffrey pines 1.2 to 1.8 m (4 to 6 ft) in diameter and 52 to 61 m (170 to 200 ft) tall were typical of the species' best growth on deep, coarse-textured and well-drained soils (28,45,52). Stands of similar description may still be seen in the high country of Yosemite National Park. By contrast, stand productivity is low for Jeffrey pine on ultramafic soils. Dunning's site index (base age 300 years) may often be as high as 29 m (95 ft), but the typical stocking capacity is just 11 to 28 percent of normal basal area (33).
Yield data have never been acquired specifically for Jeffrey pine, but Jeffrey pine apparently grows to the same age and maximum size as ponderosa pine. Observations in mature natural stands bolster the belief that yield data for ponderosa pine can be confidently applied to pure stands of Jeffrey pine, or to Jeffrey pine mixed with ponderosa pine (24,25).
Rooting Habit- Windthrow is rare for Jeffrey pine. Surveys of windthrow sales indicate that Jeffrey pine is seldom included and is highly windfirm compared with its timber associates. In juvenile through mature stages, Jeffrey pine typically has a deep taproot. The primary lateral roots are strong and extensive, some growing horizontally and others angling downward. Such root systems apparently adjust well to the physical and chemical environments encountered. In an open stand of Jeffrey pine on a shallow ultramafic soil in the northern Sierra Nevada, live roots up to 5 cm (2 in) in diameter were encountered in soil pits up to 30 m (100 ft) away from the nearest trees, at distances greater than tree height.
Reaction to Competition- Jeffrey pine occasionally regenerates beneath open, overmature stands, but growth is checked until the overstory is removed. The species is intolerant of shade, and root competition from old-growth trees is intense. In such circumstances, saplings 40 or more years old and less than 1 m (3 ft) tall are common. After release, suppressed saplings take 3 to 7 years to extend root systems, produce efficient crowns, and begin rapid height growth (24).
As a result of wildfire, stagnated sapling stands of naturally regenerated Jeffrey and ponderosa pines are common in the 1.6 million ha (4 million acres) of yellow pine forests in northeastern California. Densities have sometimes reached 42,000 stems per hectare (17,000/acre). Growth is so slow that stand development virtually ceases, yet dominants and codominants can respond to thinning. In one 55-year-old stand with 27,200 stems per hectare (11,000/acre), thinning 2.5-m (8.2-ft) tall saplings to a spacing of 2.7 m (9 ft) tripled their periodic radial growth and increased height growth 67 percent in 5 years, compared with unthinned controls (53).
Survival and growth of planted Jeffrey pine reflects the thoroughness of site preparation and post-planting protection against aggressive understory plants (42,43). Heavy invasion of any vegetation soon after planting makes seedling survival unlikely, if not impossible. Brush, grasses, and sedges all are lethal competition for available soil water in Jeffrey pine's dry summer environments, and many shrub species quickly overtop and markedly slow the growth of surviving seedlings.
Low vegetation even reduces the growth of established Jeffrey pine. In northeastern California, removing perennial bunchgrass and sedge, alone or together with sagebrush (Artemisia tridentata) and bitterbrush (Purshia tridentata), increased the mean 5-year basal area increment of pine poles by as much as 38 percent (18).
Damaging Agents- Extremely low temperatures can kill needles, buds, last season's shoots, and even the inner bark of Jeffrey pine. East of the Sierra Nevada crest, winter minima between -35° and -43° C (-31° and -45° F) have extensively damaged Jeffrey pine. Trees whose inner bark was destroyed by freezing were killed (56). Jeffrey pine may also undergo severe physiological drought and foliar damage in winter, when sunny and warm or windy days desiccate needles of trees rooted in soils that are too cold for adequate water uptake. Jeffrey pine is highly sensitive to anaerobic conditions and is killed outright when root systems are flooded by beaver ponds or new stream channels or are buried under fill from land grading operations (2).
Human activity is often damaging in other ways. Highway de-icing salts (48), sewage effluents (2), and air pollutants such as ozone (39) all may severely injure or kill Jeffrey pine. Like other conifers, Jeffrey pine is susceptible to herbicide damage at certain seasons of the year and stages of growth (40). Spraying during the growing season may kill trees or distort growth of new shoots, although observations in plantations suggest that Jeffrey pine resists phenoxy herbicide damage.
Among the biotic agents attacking Jeffrey pine are two needle diseases, a limb canker, at least five different rusts, western dwarf mistletoe, three major root diseases, and various heart rots (2).
Elytroderma disease (Elytroderma deformans) has reached epidemic proportions in stands into which cold air drains and has reduced growth and killed trees for years after major outbreaks (46). Medusa needle blight (Davisomycella medusa) has markedly decreased growth of individual trees, especially on poor sites after drought. Cenangium limb canker (Cenangium ferruginosum) may severely attack young trees growing under poor conditions and usually kills suppressed or weakened branches.
Stalactiform rust (Peridermium stalactiforme) infects lower limbs and spreads upward in the crowns of young Jeffrey pine; infected trees are almost always near the alternate hosts, particularly Castilleja spp. Filamentosum rust (Peridermium filamentosum) kills the middle or upper crowns of scattered mature trees, can spread from pine to pine, and has displayed a potential for intense outbreaks. Sweetfern rust (Cronartium comptoniae) frequently kills young trees, and tarweed rust (Coleosporium madiae) may cause heavy defoliation in occasional wet years. Western gall rust (Peridermium harknessii) kills seedlings and large trees of Jeffrey pine by producing abundant, globose branch galls or large bole-deforming stem cankers. Although its major outbreaks are often decades apart, this rust is ubiquitous in California and potentially hazardous to young Jeffrey pine in moderately cool, humid environments (2).
The worst disease of Jeffrey pine is caused by western dwarf mistletoe (Arceuthobium campylopodum). Heavy infections cause witches' brooms, severely reduce growth, and eventually kill the tree. Young trees are highly susceptible to infection from surrounding infected overstory trees (37). Dwarf mistletoe has predisposed many stands to insect attack and has induced 60 to 80 percent of all Jeffrey pine mortality in years of severe drought (4).
Fungal diseases of the roots of Jeffrey pine include annosus (Heterobasidion annosum), armillaria (Armillaria mellea), and black stain (Verticicladiella wagnerii). Fungi that cause heart rots in Jeffrey pine include species of Lentinus, Fomes, and Polyporus. In southern California, red rot (Dichomitus squalens) attacks Jeffrey pine through broken tops, bole wounds, and large dead limbs (2).
Insects that damage Jeffrey pine are as numerous as the fungal diseases. Collectively attacking every part of the tree, they include twig and needle scales, various defoliators, borers, and tip moths, several bark beetles, and a host of cone and seed feeders (17).
The ponderosa pine twig scale (Matsucoccus bisetosus) feeds on branches and stems of trees of all ages and is the most destructive of the scales. Two of the more serious defoliators are the pine needle sheathminer (Zelleria haimbachi) and pandora moth (Coloradia pandora). Larvae of the sheathminer have destroyed more than 75 percent of the new needles in localized outbreaks in California. Larvae of the pandora moth are among the largest of any forest insect and consume whole needles in spring before bud break. Extensive outbreaks occur every 20 to 30 years, cause heavy defoliation for 2 to 4 years, and predispose the trees to attack by bark beetles and borers.
The western pineshoot borer (Eucosma sonomana) stunts the needles and retards height growth of young trees by as much as 30 percent annually. The ponderosa pine tip moth (Rhyacionia zozana) kills current shoots of saplings and young trees up to 2 m (6 ft) tall and may chronically retard growth for many years. The fir coneworm (Dioryctria abietivorella) kills the terminal buds of saplings and poles and frequently causes a permanent fork in the main stem. Larvae of the pine reproduction weevil (Cylindrocopturus eatoni) can destroy saplings and young trees where brush competition causes severe water stress.
The Jeffrey pine beetle (Dendroctonus jeffreyi) is the single worst enemy of Jeffrey pine (13). This bark beetle is prevalent throughout the range of its host, and has caused staggering losses of timber in mature stands. Several other insects that attack the main stem commonly precede or accompany Jeffrey pine beetle, notably the California flatheaded borer (Melanophila californica) and two pine engravers, the emarginate ips (Ips emarginatus) and Oregon pine engraver (I. pini, or I. oregonis). The red turpentine beetle (Dendroctonus valens) also attacks injured or weakened Jeffrey pine and induces fatal attacks by other bark beetles. Throughout California, lethal bark beetle activity is usually associated with dwarf mistletoe infestation or root rots, indicating that pathogens predispose the trees to insect attacks (4).
Insects often cause major losses of Jeffrey pine cones and seeds. The worst of these are the Jeffrey pine seedworm (Laspeyresia injectiva) and ponderosa pine seedworm (L. piperana), which eat seeds within the immature cone (17,26).
Deer, jack rabbits and snowshoe hares, pocket gophers, porcupines, and domestic livestock damage and kill young Jeffrey pine. Resident, mobile populations of these mammals make substantial losses likely in most areas. Pocket gophers consume whole seedlings, feed on the roots, stem, and crown of saplings, and often annihilate young plantations (11). Porcupines commonly eat the inner bark and cambium of saplings and poles and either kill them outright or cause a spiketop above the girdled stem.
Special Uses
In commerce, no distinction is made between the wood of Jeffrey pine and that of ponderosa pine.
Genetics
Jeffrey pine is genetically variable. Estimates of the average number of alleles and average heterozygosity per enzyme locus show its allelic variation is high (7).
Population Differences
Jeffrey pine displays large differences in seedling and sapling growth, and in most nurseries seed source largely determines seedling size. When raised in a nursery in the western Sierra Nevada, dormant seedlings of Jeffrey pine from east of the Sierra Nevada crest develop top growth capacity earlier in winter than sources from the western Sierra and southern California. At the same time, seedlings of all sources from east of the crest and southern California show two separate peaks in root growth capacity: November and January for the eastern Sierra, and December and February or March for southern California. The depression between peaks can be deep or shallow and span 1 or 2 months. By contrast, seedlings from ultramafic soil in the western Sierra show a single peak in midwinter (27).
Growth in young plantations markedly depends on seed source. In the North Coast Range, stem volumes of 20-year-old Jeffrey pine from five sources in the northern Sierra Nevada differed by up to twofold (6). Trees with least growth were from seed parents near 2230 m (7,300 ft), and those with most growth from parents on ultramafic soil near 1050 m (3,450 ft). Trees from parents at the highest elevation, at 2510 m (8,235 ft) near the Sierra crest, were notably larger than trees from 1770, 1920, and 2230 m (5,800, 6,300, and 7,300 ft).
In the western Sierra Nevada, performance of 20-year-old Jeffrey pine improved with the match between seed source and planting site elevation (5). At low elevations, trees from 1050 m (3,450 ft) were taller and had less stem taper than trees from 2400 m (7,860 ft). At high elevations, mortality was 30 percent in the low elevation source, as against 2 percent in high elevation sources. Trees of high elevation sources were 1. 1 to 1.6 m (3.6 to 5.2 ft) taller, and fewer than 10 percent suffered breaks in the main stem, compared with 27 percent in the low elevation source.
Data at hand are not sufficient to describe the rangewide pattern of genetic variation in the growth of Jeffrey pine. Definitive data are probably years away, because early growth of Jeffrey pine depends on specific interactions between the seed source and planting site environment. Until rotation-age tests of particular sources on specific sites indicate otherwise, planting any Jeffrey pine of a nonlocal source is unwise. Nonlocal sources are probably maladapted, yet may not show it until sometime during or after the pole stage.
There is good evidence that some Jeffrey pine may be resistant to dwarf mistletoe infection (47,57).
Hybrids
In certain stands, Jeffrey pine is known to hybridize with ponderosa pine (23) and Coulter pine (60), but most mixed stands contain no morphological evidence of genetic intermingling. Natural hybrids are rare because these pines differ substantially in flowering time (12) and cross only with difficulty (8).
Under controlled conditions, Jeffrey pine will cross with either ponderosa pine or Coulter pine (32,41), but crossability is very low in either combination. With Jeffrey pine as the female parent, 40 percent of the strobili dusted with ponderosa pollen may produce cones, and 3 percent of the seeds may be sound; with Coulter pollen, the expected percentages are 61 and 3 (8). Observed crossing barriers include failure of viable pollen to germinate on the nucellus, failure of pollen tubes to maintain normal growth rates in the nucellus, lack of exact timing between tube growth and egg formation, collapse of the proembryo, and failure of the embryo to develop. In both crosses, slow growth of the pollen tube appears to be the most common barrier to development of hybrid seeds (29).
Although hybrids of Jeffrey and ponderosa pines are rare in the upper Mixed Conifer Forest and in sympatric distributions east of the Sierra-Cascade crest, hybrids may be locally common on ultramafic soils in the western Sierra Nevada. Of 13 mature Jeffrey pines sampled on an ultramafic soil in Sierra County, CA, 12 had the expected 95 to 99 percent heptane, but one had 75 percent heptane and 7 to 8 percent each of alpha-pinene, beta-pinene, and limonene, which indicate ponderosa pine ancestry (50). Beneath overstories of Jeffrey pine on ultramafic soil in Placer County, CA, individual saplings and poles show a diverse mix of morphological traits, suggesting that the pollen parents were ponderosa pines growing on the same soil or in mixed conifer forests nearby.
Capitalizing on Coulter pine's immunity to the pine reproduction weevil, the Forest Service produced backcrosses of Jeffrey-Coulter hybrids to Jeffrey pine on a large scale for many years (8). The weevil is controllable silviculturally, however, and in recent years the demand for backcross seedlings has ceased.
Literature Cited
- Axelrod, Daniel 1. 1976. History of the coniferous forests, California and Nevada. University of California Publications in Botany 70:1-61.
- Bega, Robert V., tech. coord. 1978. Diseases of Pacific Coast conifers. U.S. Department of Agriculture, Agriculture Handbook 521. Washington, DC. 206 p.
- Billings, W. D. 1950. Vegetation and plant growth as affected by chemically altered rocks in the western Great Basin. Ecology 31:62-74.
- Byler, James W. 1978. The pest damage inventory in California. In Proceedings, Symposium on Dwarf Mistletoe Control Through Forest Management, April 11-13, 1978, Berkeley, CA. p. 162-171. USDA Forest Service, General Technical Report PSW-31. Pacific Southwest Forest and Range Experiment Station, Berkeley, CA.
- Callaham, R. Z., and A. R. Liddicoet. 1961. Altitudinal variation at 20 years in ponderosa and Jeffrey pines. Journal of Forestry 59:814-820.
- Callaham, R. Z., and Woodbridge Metcalf. 1959. Altitudinal races of Pinus ponderosa confirmed. Journal of Forestry 57:500-502.
- Conkle, M. Thompson. 1981. Isozyme variation and linkage in six conifer species. In Proceedings, Symposium on Isozymes of North American Forest Trees and Forest Insects, July 27, 1979, Berkeley, CA. p. 11-17. USDA Forest Service, General Technical Report PSW-48. Pacific Southwest Forest and Range Experiment Station, Berkeley, CA.
- Critchfield, William B. 1966. Crossability and relationships of the California big-cone pines. In USDA Forest Service, Research Paper NC-6. p. 36-44. North Central Forest Experiment Station, St. Paul, MN.
- Critchfield, William B. 1971. Profiles of California vegetation. USDA Forest Service, Research Paper PSW-76. Pacific Southwest Forest and Range Experiment Station, Berkeley, CA. 54 p.
- Critchfield, William B., and Elbert L. Little, Jr. 1966. Geographic distribution of the pines of the world. U.S. Department of Agriculture, Miscellaneous Publication 991. Washington, DC. 97 p.
- Crouch, Glenn L. 1971. Susceptibility of ponderosa, Jeffrey, and lodgepole pines to pocket gophers. Northwest Science 45:252-256.
- Duffield, J. W. 1953. Pine pollen collection dates-annual and geographic variation. USDA Forest Service, Research Note 85. California Forest and Range Experiment Station, Berkeley. 9 p,
- Eaton, Charles B. 1956. Jeffrey pine beetle. U.S. Department of Agriculture, Forest Pest Leaflet 11. Washington, DC. 7 p.
- Eyre, F. H., ed. 1980. Forest cover types of the United States and Canada. Society of American Foresters, Washington, DC. 148 p.
- Fowells, H. A. 1953. The effect of seed and stock sizes on survival and early growth of ponderosa and Jeffrey pine. Journal of Forestry 51:504-507.
- Fowells, H. A., and N. B. Stark. 1965. Natural regeneration in relation to environment in the mixed conifer forest type of California. USDA Forest Service, Research Paper PSW-24. Pacific Southwest Forest and Range Experiment Station, Berkeley, CA. 14 p.
- Furniss, R. L., and V. M. Carolin. 1977. Western forest insects. U.S, Department of Agriculture, Miscellaneous Publication 1339. Washington, DC. 654 p.
- Gordon, Donald T. 1962. Growth response of east side pine poles to removal of low vegetation. USDA Forest Service, Research Note 209. Pacific Southwest Forest and Range Experiment Station, Berkeley. 3 p.
- Griffin, James R. 1974. A strange forest in San Benito County. Fremontia 2:11-15.
- Griffin, James R., and William B. Critchfield. 1972. The distribution of forest trees in California. USDA Forest Service, Research Paper PSW-82. (Rev. with suppl. 1976). Pacific Southwest Forest and Range Experiment Station, Berkeley, CA. 118 p.
- Haller, John R. 1959. Factors affecting the distribution of ponderosa and Jeffrey pines in California. Madroño 15:65-71.
- Haller, John R. 1961. Some recent observations on ponderosa, Jeffrey and Washoe pines in northeastern California. Madroño 16:126-132.
- Haller, John R. 1962. Variation and hybridization in ponderosa and Jeffrey pines. University of California Publications in Botany 34:123-166.
- Hallin, William E. 1957. Silvical characteristics of Jeffrey pine. USDA Forest Service, Technical Paper 17. California Forest and Range Experiment Station, Berkeley. 11 p.
- Hallin, William E. 1959. The application of unit area control in the management of ponderosa-Jeffrey pine at Blacks Mountain Experimental Forest. U.S. Department of Agriculture, Technical Bulletin 1191. Washington, DC. 96 p.
- Hedlin, Alan F., Harry 0. Yates III, David Cibrian Tovar, and others. 1980. Cone and seed insects of North American conifers. USDA Forest Service, Washington, DC. 122 p.
- Jenkinson, James L. 1980. Improving plantation establishment by optimizing growth capacity and planting time of western yellow pines. USDA Forest Service, Research Paper PSW-154. Pacific Southwest Forest and Range Experiment Station, Berkeley, CA. 22 p.
- Jepson, Willis Linn. 1923. The trees of California. University of California, Berkeley. 240 p.
- Krugman, Stanley L. 1970. Incompatibility and inviability systems among some western North American pines. In Proceedings, International Union of Forestry Research Organizations, Section 22, Working Group, Part II, Sexual reproduction of forest trees. IUFRO, Finland. 13 p.
- Krugman, Stanley L., and James L. Jenkinson. 1974. Pinus L. pine. In Seeds of woody plants in the United States. C. S. Schopmeyer, tech. coord. p. 598-638. U.S. Department of Agriculture, Agriculture Handbook 450. Washington, DC.
- Laacke, Robert J. 1979. California forest soils. University of California Division of Agricultural Sciences, Priced Publication 4094. Berkeley. 181 p.
- Little, Elbert L., Jr., and Francis I. Righter. 1965. Botanical descriptions of forty artificial pine hybrids. U.S. Department of Agriculture, Technical Bulletin 1345. Washington, DC. 47 p.
- MacLean, Colin D., and Charles L. Bolsinger. 1973. Estimating productivity on sites with a low stocking capacity. USDA Forest Service, Research Paper PNW-152. Pacific Northwest Forest and Range Experiment Station, Portland, OR. 18 P.
- Oliver, William W. 1972. Growth after thinning ponderosa and Jeffrey pine pole stands in northeastern California. USDA Forest Service, Research Paper PSW-85. Pacific Southwest Forest and Range Experiment Station, Berkeley, CA. 8 p.
- Oliver, William W. 1979. Fifteen-year growth patterns after thinning a ponderosa-Jeffrey pine plantation in northeastern California. USDA Forest Service, Research Paper PSW-141. Pacific Southwest Forest and Range Experiment Station, Berkeley, CA. 10 p.
- Pardo, Richard. 1978. National register of big trees. American Forests 84(4):17-47.
- Parmeter, J. R., and Robert F. Scharpf. 1972. Spread of dwarf mistletoe from discrete seed sources into young stands of ponderosa and Jeffrey pines. USDA Forest Service, Research Note PSW-269. Pacific Southwest Forest and Range Experiment Station, Berkeley, CA. 5 p,
- Peterson, Glenn W., and Richard S. Smith, Jr., tech. coords. 1975. Forest nursery diseases in the United States. U.S. Department of Agriculture, Agriculture Handbook 470. Washington, DC. 125 p.
- Pronos, John, Detlev R. Vogler, and Richard S. Smith, Jr. 1978. An evaluation of ozone injury to pines in the southern Sierra Nevada. USDA Forest Service, Forest Insect and Disease Management Report 78-1. San Francisco, CA. 17 p.
- Radosevich, S. R., E. J. Roncoroni, S. G. Conard, and W. B. McHenry. 1980. Seasonal tolerance of six coniferous species to eight foliage-active herbicides. Forest Science 26:3-9.
- Righter, F. I., and J. W. Duffield. 1951. Interspecies hybrids in pines. Journal of Heredity 42:75-80.
- Roy, D. F. 1953. Effects of ground cover and class of planting stock on survival of transplants in the eastside pine type of California. USDA Forest Service, Research Note 87. California Forest and Range Experiment Station, Berkeley. 6 p.
- Roy, Douglass F. 1981. Effects of competing vegetation on conifer performance. Paper presented at Forest Vegetation Workshop, March 3-5, 1981, School of Forestry, Oregon State University, Corvallis. Pacific Southwest Forest and Range Experiment Station, Berkeley, CA. 54 p.
- Rundel, Philip W., David J. Parsons, and Donald T. Gordon. 1977. Montane and subalpine vegetation of the Sierra Nevada and Cascade Ranges. In Terrestrial vegetation of California. p. 559-599. Michael G. Barbour and Jack Major, eds. John Wiley, New York.
- Sargent, Charles Sprague. 1965. Manual of the trees of North America. vol. 1. Dover, New York. 433 p.
- Scharpf, Robert F., and Robert V. Bega. 1981. Elytroderma disease reduces growth and vigor, increases mortality of Jeffrey pines at Lake Tahoe Basin, California. USDA Forest Service, Research Paper PSW-155. Pacific Southwest Forest and Range Experiment Station, Berkeley, CA. 6 p.
- Scharpf, Robert F., and J. R. Parmeter, Jr. 1967. Spread of dwarf mistletoe into Jeffrey pine plantation ... trees infected after 22 years. USDA Forest Service, Research Note PSW-141. Pacific Southwest Forest and Range Experiment Station, Berkeley, CA. 6 p.
- Scharpf, Robert F., and Michael Srago. 1974. Conifer damage and death associated with the use of hiway deicing salt in the Lake Tahoe Basin of California and Nevada. USDA Forest Service, California Region Forest Pest Control Technical Report 1. San Francisco, CA. 16 p.
- Siggins, Howard W. 1933. Distribution and rate of fall of conifer seeds. Journal of Agricultural Research 47:119-128.
- Smith, Richard H. 1967. Monoterpene composition of pine species and hybrids ... some preliminary findings. USDA Forest Service, Research Note PSW-135. Pacific Southwest Forest and Range Experiment Station, Berkeley, CA. 14 p.
- Stone, Edward C. 1957. Dew as an ecological factor. II. The effect of artificial dew on the survival of Pinus ponderosa and associated species. Ecology 38:414-422.
- Sudworth, George B. 1967. Forest trees of the Pacific Slope. Dover, New York. 455 p.
- Teeguarden, Dennis E., and Donald T. Gordon. 1964. Thinning increases growth of stagnated ponderosa and Jeffrey pine sapling stands. Journal of Forestry 62:114.
- Tomback, Diana F. 1978. Foraging strategies of Clark's nutcracker. The Living Bird 16:123-161.
- Vasek, Frank C. 1978. Jeffrey pine and vegetation of the southern Modoc National Forest. Madroño 25:9-30.
- Wagener, Willis W. 1960. A comment on cold susceptibility of ponderosa and Jeffrey pines. Madroño 15:217-219.
- Wagener, Willis W. 1965. Dwarf mistletoe removal and reinvasion in Jeffrey and ponderosa pine, northeastern California. USDA Forest Service, Research Note PSW-73. Pacific Southwest Forest and Range Experiment Station, Berkeley, CA. 8 p.
- Waring, R. H. 1969. Forest plants of the Eastern Siskiyous: their environment and vegetational distribution. Northwest Science 43:1-17.
- Waring, R. H., and J. Major. 1964. Some vegetation of the California coastal redwood region in relation to gradients of moisture, nutrients, light, and temperature. Ecological Monographs 34:167-215.
- Zobel, Bruce. 1951. The natural hybrid between Coulter and Jeffrey pines. Evolution 5:405-418,