
Abies balsamea (L.) Mill.
Balsam Fir
Pinaceae -- Pine family
By: Robert M. Frank
From: Burns, Russell M., and Barbara H. Honkala, tech. coords. 1990. Silvics of North America: 1. Conifers; Agriculture Handbook 654. U.S. Department of Agriculture, Forest Service, Washington, DC. vol.2, 877 p.
Balsam fir (Abies balsamea) is one of the more important conifers in the northern United States and in Canada. Within its range it may also be referred to as balsam, Canadian balsam, eastern fir, and bracted balsam fir. It is a small to medium-sized tree used primarily for pulp and light frame construction, and it is one of the most popular Christmas trees. Wildlife rely extensively on this tree for food and shelter.
Habitat
Native Range
In Canada, balsam fir extends from Newfoundland and Labrador west through the more northerly portions of Quebec and Ontario, in scattered stands through north-central Manitoba and Saskatchewan to the Peace River Valley in northwestern Alberta, then south for approximately 640 km (400 mi) to central Alberta, and east and south to southern Manitoba.
In the United States, the range of balsam fir extends from extreme northern Minnesota west of Lake-of-the-Woods southeast to Iowa; east to central Wisconsin and central Michigan into New York and central Pennsylvania; then northeastward from Connecticut to the other New England States. The species is also present locally in the mountains of Virginia and West Virginia (23,30).
Balsam fir grows from sea level to within 15 to 23 m (50 to 75 ft) below the 1917 m (6,288 ft) summit of Mount Washington in the White Mountains of New Hampshire. At this elevation prostrate balsam fir is found in sheltered areas (1).
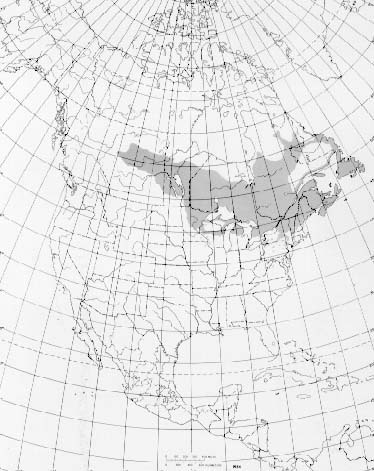
- The native range of balsam fir.
Climate
Balsam fir grows best in the eastern part of its range in southeastern Canada and the Northeastern United States. This area is characterized by cool temperatures and abundant moisture. Growth is optimum in areas with a mean temperature of 2° to 4° C (35° to 40° F), a January average ranging from -18° to -12° C (0° to 10° F), a July mean temperature ranging from 16° to 18° C (60° to 65° F), and mean annual precipitation ranging from 760 to 1100 mm (30 to 43 in) (1).
The mean annual temperature within the range of balsam fir varies from -4° to 7° C (25° to 45° F). Mean annual precipitation records show as much as 1400 mm (55 in) to as little as 390 mm (15 in). The amount of growing season precipitation is from 150 to 620 mm (6 to 25 in) (1). There are 80 to 180 frost-free days and about 110 days for optimum growth (1).
Soils and Topography
Balsam fir grows on a wide range of inorganic and organic soils originating from glaciation and generally falling within the acid Spodosol, Inceptisol, and Histosol soil orders. These are characterized by a thick mor humus and a well-defined A2 horizon, usually gray in appearance because of leaching, and commonly caused by abundant rainfall, cool climate, and coniferous cover. Many of the glacial till soils in New England are shallow and have a compact layer about 46 cm (18 in) below the surface (11).
Soil moisture was the most important predictor of site index in a study in Newfoundland. Soil nutrient status and topography, in that order, were of lesser importance. Glacial tills, often shallow, cover much of the area (27).
Balsam fir has been reported as growing on soils of a wide range of acidity. In the northern Lake States it is most common on cool, wet-mesic sites with pH values between 5.1 to 6.0 (19). Optimum growth occurs on soils where the pH of the upper organic layers is between 6.5 and 7.0 (1). On gravelly sands and in peat swamps, growth is comparatively slow (41).
Associated Forest Cover
Tree species associated with balsam fir in the boreal region of Canada are black spruce (Picea mariana), white spruce (Picea glauca), paper birch (Betula papyrifera), and quaking aspen (Populus tremuloides). In the more southerly northern forest region, additional associates include bigtooth aspen (Populus grandidentata), yellow birch (Betula alleghaniensis), American beech (Fagus grandifolia), red maple (Acer rubrum), sugar maple (Acer saccharum), eastern hemlock (Tsuga canadensis), eastern white pine (Pinus strobus), tamarack (Larix laricina), black ash (Fraxinus nigra), and northern white-cedar (Thuja occidentalis). Red spruce (Picea rubens) is an important associate in New Brunswick and Maine. Occasional associates are balsam poplar (Populus balsamifera), gray birch (Betula populifolia), red pine (Pinus resinosa), jack pine (Pinus banksiana), and American elm (Ulmus americana) (10).
Pure stands of balsam fir or stands in which balsam fir is the major component of growing stock make up the forest cover type Balsam Fir (Society of American Foresters Type 5) (10). Balsam fir is also a major component in two other eastern forest cover types: Red Spruce-Balsam Fir (Type 33) and Paper Birch-Red Spruce-Balsam Fir (Type 35). It is an associated species in 22 eastern forest cover types and in 4 western forest cover types.
Common shrubs associated with balsam fir include beaked hazel (Corylus cornuta), mountain maple (Acer spicatum), Labrador-tea (Ledum groenlandicum), Canada yew (Taxus canadensis), red raspberry (Rubus idaeus var. strigosus), sheep-laurel (Kalmia angustifolia), and hobblebush (Viburnum lantanoides) (10,41).
Among the herbaceous plants commonly found under balsam fir are twinflower (Linnaea borealis), bunchberry (Cornus canadensis), starflower (Trientalis borealis), creeping snowberry (Gaultheria hispidula), sedges (Carex spp.), common woodsorrel (Oxalis montana), bluebead lily or cornlily (Clintonia borealis), painted trillium (Trillium undulatum), cinnamon fern (Osmunda cinnamomea), sweetscented bedstraw (Galium triflorum), Canada mayflower (Maianthemum canadense), and spinulose woodfern (Dryopteris spinulosa).
Certain associations of shrubs, herbs, and mosses indicate forest site quality (41). The four main indicator associations, designated as Hylocomium/ Hypnum, Cornus/Maianthemum, Oxalis/Cornus, and Viburnum/Oxalis indicate, in the order listed, increasing productivity of site and increasing proportions of shrubs and hardwood trees in natural stands. Only the Hylocomium/Hypnum sites are likely to be occupied by pure balsam fir.
Life History
Reproduction and Early Growth
Flowering and Fruiting- Exposure to light influences flowering in balsam fir. In New Brunswick, female strobili were observed on 83 percent of dominant, 59 percent of codominant, and 6 percent of intermediate trees. None were found on suppressed trees (41).
Balsam fir is monoecious. In spring, 1 year before pollination, male (staminate) and female (ovulate or pistillate) strobili differentiate from flower buds. The strobili are microscopically recognizable at this time. Male strobili usually are distinguishable before the female strobili because they initially develop more rapidly. Flower buds usually open in late May or early June before vegetative buds (41) but have been reported as flowering as early as late April (42).
Male strobili, yellowish-red and tinged with purple, develop in the axils of leaves along the undersides of the 1-year-old twigs, usually in dense clusters. Their position in the crown is mostly within 5 m (15 ft) of the top and is almost always below the female strobili. Female strobili are purplish and are found singly or in small groups, confined to the top 1.5 m (5 ft) of the crown. They are located on the upper side of the twig and, like the male strobili, develop on the previous year's twig. Flower production is best on the outer end of branches (41,42). At maturity, male flowers are about 3 mm (0.1 in) long; female flowers are about 25 mm (1.0 in) long (1).
Pollen grains are yellow; when developed, their average diameter is 90 µ (0.00354 in). In one series of observations in Ontario, fertilization occurred on June 25 (1). The mature fruit is an erect cone 5 to 10 cm (2 to 4 in) long with short, round, irregularly notched scales and pointed tips. There are thin, closely overlapping fan-shaped scales near the center of the cone. The cone matures and ripens during the first fall in late August and early September. The scales and shorter bracts drop away with the seeds, leaving the central axis, which can persist for many years.
Seed Production and Dissemination- Regular seed production probably begins after 20 to 30 years. Cone development has been reported for trees 15 years of age and younger and only 2 m (6.6 ft) tall. Good seed crops occur at intervals of 2 to 4 years, with some seed production usually occurring during intervening years (1). On the average, 35 L (bushel) containing 1,000 to 2,000 cones weighs approximately 16 kg (35 lb) and yields 1000 to 1200 g (35 to 42 oz) of cleaned seeds. The number of cleaned seeds per kilogram (2.2 lb) ranges from 66,000 to 208,000 and averages 131,000. These are about 134 seeds per cone (42). The seed yield of balsam fir ranged from 5.6 to 20.2 kg/ha (5 to 18 lb/acre) during several good seed years in Ontario (1). Over a 37-year period, annual seed production in this area averaged 1,950 seeds per square meter (181/ft²) (15).
The period of balsam fir seedfall is long and dissemination distances vary. Seedfall begins late in August, peaks in September and October, and continues into November. Some seeds fall throughout the winter and into early spring. Most of the seeds are spread by wind-some to great distances over frozen snow-and some are spread by rodents. Although seeds may disseminate from 100 m (330 ft) to more than 160 m (525 ft), effective distances are 25 m to 60 m (80 to 200 ft) (1,11,28). Many seeds falling with the cone scales land close to the base of the tree.
Balsam fir seeds have dormant embryos and should be stratified in moist sand at about 50 C (410 F) for at least 30 days before planting. Germination is epigeal (42).
Seedling Development- Within the range of suitable temperatures, moisture is more important than light for germination. In fact, light intensities of only 10 percent of full sunlight result in successful germination (1). The low capacity of planted balsam fir seeds to germinate may be attributed in part to seed injury during the cleaning process. The age of the tree may also contribute to the viability of seeds.
A study in Michigan (41) showed that germination was highest for a 41-year-old tree (68 percent), varied for trees 30 years old (8 to 57 percent), and was lowest for trees 155 years old (10 percent). Testing of 32 commercial seed lots showed average germination of about 26 percent with a range of 4 to 62 percent (42). Once the seed reaches the ground, its viability diminishes quickly and is gone within 1 year (13). It has been suggested, however, that in cold swamps viability of some seeds is retained for 2 to 3 years (1).
Most germination occurs from late May to early July. Survival the first winter is questionable if germination occurs after mid-July (1). If enough moisture is available, almost any seedbed type is satisfactory, but mineral soil-neither too sandy nor too heavy-with some shade is best. Litter and humus are poor seedbeds, especially if moisture is inadequate or -light is excessive. Competition, often severe, makes heavy sod the poorest seedbed (11).
A thick layer of duff exceeding about 8 cm (3 in) is less favorable for balsam fir but even worse for the slower growing associated spruces. Balsam fir seedlings may have a heavy central root, much like a taproot, that extends to the bottom of the humus layer and then splits into several laterals. In general, balsam fir roots grow more rapidly and penetrate deeper than red spruce roots. Where seasonal root elongation of young balsam fir growing in humus averaged 10.6 cm (4.2 in), red spruce was 7.6 cm (3.0 in), and white spruce 9.0 cm (3.5 in), or 39 percent and 18 percent less, respectively (1).
Because the surface of thick duff usually dries out, there may be some delayed germination as late as August. Few seedlings become established, however. The closer seeds lie to mineral soil, the greater the initial establishment of seedlings.
Seedlings starting in the open may sustain heavy mortality when surface temperatures exceed 46° to 54° C (115° to 130° F) or when there is drought or frost heaving. Seedlings may also be smothered or crushed by litter, ice, snow, and hardwood leaves. Losses after the first year usually are minor. As seedlings develop, light at intensities of at least 50 percent of full sunlight are necessary for optimum growth (11,41). Damage caused by late spring frost to new foliage of young seedlings is seldom severe.
Balsam fir seedlings about 15 cm (6 in) tall can be considered to be established (11), especially if secondary branching has occurred. Early growth is then determined largely by the amount and character of dominant competition. Bracken, raspberry, and hardwood sprouts-especially the maples-are the chief competitors on heavily cutover lands in the Northeast. These species may increase dramatically when the original basal area is reduced by 50 percent or more and may dominate the site for 10 to 25 years (2). Unless there has been some soil disturbance, there will be little regeneration of balsam fir and spruce immediately following logging (45). Both balsam fir and the spruces can survive many years of suppression and still respond to release (11,41). The space required for the continual development and establishment of new seedlings probably exceeds that created by the removal of individual trees. To ensure successful regeneration relatively small groups of trees should be removed initially (12).
Vegetative Reproduction- Layering is not an important means of regeneration except for prostrate balsam fir growing in the more northern and mountainous locations such as Isle Royale in Lake Superior, and the White Mountains of New Hampshire. Layering also occurs in open swamps and deep mossy areas and under white pine and jack pine overstories. Trees of any age apparently may layer. Second generations, vegetatively produced, develop when connecting tissues decay and separate (1).
Balsam fir apparently grafts easily (41). In a study in New York, greenhouse grafts were 85 percent successful and field grafts were 80 percent successful. One attempt to air-layer balsam fir was unsuccessful (1). Balsam fir Christmas trees are stump cultured from lateral branches or adventitious shoots.
Sapling and Pole Stages to Maturity
Growth and Yield- Balsam fir at maturity is small to medium size, depending on location and growing conditions. In general, heights range from 12 to 18 m (40 to 60 ft); diameters range from 30 to 46 cm. (12 to 18 in) at breast height (41). Where growth is optimum, as in the Green River watershed in New Brunswick, some trees can reach 27 m (90 ft) in height and 75 cm. (30 in) in d.b.h. The reported record d.b.h. for balsam fir is 86 cm (34 in). Maximum age is about 200 years (1). How large or how fast balsam fir grows, or how much a stand of balsam fir will yield is related to site factors such as biotic, climatic, and soil conditions, and to age. The condition of the tree or stand and the composition and structure of the stand also influence growth.
Diameter growth was related to vigor and crown length-to-height ratio in a study in Maine. Balsam fir with high vigor and a ratio of at least 0.7- the proportion of live-crown length to total tree height averaged 6.1 cm (2.4 in) of growth in d.b.h. in 10 years. Less vigorous trees with smaller crown-length ratios ranged downward to an average of 1.0 cm (0.4 in) of growth in 10 years. Vigorous trees with room to grow attain a d.b.h. of at least 25 cm (10 in) in about 50 years (41). In uneven-aged stands of several density classes in Maine, balsam fir grew faster in diameter than spruce and hemlock (35).
Data obtained from stem analysis of balsam fir growing on sites of varying quality in northern Maine has shown height growth curves to be polymorphic (fig. 1). Height growth varies with site quality. From these curves the average site index of a stand can be estimated (16). Monomorphic or harmonized site index curves for balsam fir are also available (17).
Balsam fir is a strong contender for space in stands in which it grows. A 20-year record of stands containing balsam fir in the Penobscot Experimental Forest in Maine showed that the periodic annual volume ingrowth of the species, as a proportion of total volume ingrowth, greatly exceeded its representation in the original stands (12). Because of its many natural enemies, however, volume mortality of balsam fir also greatly exceeds its original representation in these stands.
Balsam fir accounted for 35 percent of the average annual net growth in predominantly softwood stands and 32 percent in mixed stands that were extensively managed. These stands were growing at annual rates of 3.5 m³/ha (49.3 ft³/acre) and 2.9 m³/ha (41.1 ft³/acre), respectively (31).
Yields in total cubic-foot volume, including stump and top, of all trees larger than 1.5 cm (0.6 in), in d.b.h. are given in table 1. These yields are based on sample plots in even-aged spruce-fir stands, mostly on old fields. They tend to exaggerate the yields that might be expected from the irregular stands that develop after harvesting (41).
| Site index¹ | ||||
| 12.2 m
or 40 ft |
15.2 m
or 50 ft |
18.3 m
or 60 ft |
21.3 m
or 70 ft |
|
| Age | ||||
| yr | m³/ha | |||
| 20 | 6 | 8 | 9 | 12 |
| 30 | 50 | 67 | 85 | 102 |
| 40 | 136 | 182 | 229 | 276 |
| 50 | 204 | 274 | 344 | 414 |
| 60 | 245 | 329 | 413 | 497 |
| 70 | 267 | 360 | 452 | 543 |
| 80 | 286 | 384 | 481 | 579 |
| 90 | 300 | 403 | 506 | 609 |
| yr | ft³/acre | |||
| 20 | 80 | 110 | 135 | 165 |
| 30 | 720 | 960 | 1,210 | 1,455 |
| 40 | 1,940 | 2,600 | 3,270 | 3,940 |
| 50 | 2,190 | 3,920 | 4,920 | 5,910 |
| 60 | 3,500 | 4,700 | 5,900 | 7,100 |
| 70 | 3,820 | 5,140 | 6,450 | 7,760 |
| 80 | 4,080 | 5,480 | 6,870 | 8,270 |
| 90 | 4,290 | 5,760 | 7,230 | 8,700 |
¹Base age 50 years when age is measured at d.b.h.- total tree age is estimated to be 65 years at that time. |
||||
Simulating the management and growth of forest stands containing balsam fir is possible because of advances in computer technology. A matrix model, FIBER (36), has been developed for stands in the Northeast. Even-aged and multi-aged stands, containing balsam fir, spruce, northern hardwoods, and other associated species, can be programmed to simulate a range of silvicultural treatments.
In a ranking with both hardwoods and softwoods from around the world, balsam fir is highest with a total above-ground ovendry biomass at age 50 of 184 t/ha (82 tons/acre). Annual increment or annual net primary production averages 10.3 t/ha (4.6 tons/acre) (20). In New Brunswick (3), dry-matter production of balsam fir in pure stands increased dramatically with increases in stand densities of from 1,730 stems per hectare (700/acre) to 12,350/ha (5,000/acre). At an average age from release of 43 years, total above-ground biomass was 96 t/ha (43 tons/acre) for the least dense stand and 143 t/ha (64 tons/acre) for the most dense stand.
Rooting Habit- Balsam fir root systems are mostly confined to the duff layer and to the upper few centimeters of mineral soil (11). Windfall potential is high. Damage from wind is especially likely when the shallow root systems are loosened by heavy rainfall and gusty winds and where timber removals from stands not previously thinned have been poorly conducted. These usually older, dense stands are susceptible probably because root development has been poor.
Root penetration on deep or shallow soils extends to 60 to 75 cm (24 to 30 in) and has been reported to a depth of 137 cm (54 in) in sandy soils in northern Ontario. Lateral roots of balsam fir are usually strongly developed and extend horizontally in all directions to 1.5 m (5 ft) or more (1).
Root breakage and other root damage caused by swaying trees may not be as severe as is commonly thought. Most investigators agree, however, that some root breakage probably occurs because of frostheaving and swaying. During epidemics of spruce budworm (Choristoneura fumiferana), rootlet mortality can reach 75 percent after 3 consecutive years of defoliation (1).
Balsam fir root grafts are probably common and have been reported frequently. Abrasion of the bark of roots of swaying trees on lowland soils and interroot compatibility and growth pressure on upland soils apparently account for the majority of root grafts. Infection may spread through grafted roots to damage other balsam fir trees (1).
Reaction to Competition- Balsam fir has a strong ability to become established and grow under the shade of larger trees (7,11). It is classified as very tolerant. Because relative tolerance of species may vary with soil fertility, climate, and age, balsam fir is rated as both more and less shade tolerant than red spruce, and more tolerant than either black or white spruce (41). Intraspecific competition is evident in many sapling and small pole-size stands of pure balsam fir. As these stands mature, dominance usually is expressed. Competition is severe in dense fir thickets, however, and growth rates of individual trees suffer greatly. Other major competition is from the shade-tolerant hardwoods.
In New England, balsam fir is considered a subclimax type, except that it may be a climax species in the zone below timberline. It tends to become climax in Quebec and in the Lake States (41).
Damaging Agents- Many agents act to hinder the growth of balsam fir. Insects and diseases may be devastating. Flammable needles, often close to the ground, shallow root systems, and thin resinous bark make balsam fir susceptible to severe damage and mortality from fire. Susceptibility to wind damage is especially high in old unmanaged stands growing on wet shallow soils. Various species of mice, voles, and birds consume balsam fir seed; birds and squirrels nip buds; and black bears girdle mature trees.
Balsam fir has several insect enemies, the most important by far being the spruce budworm. Despite its name, the spruce budworm prefers fir over spruce; it is most likely to cause heavy damage and mortality in stands that contain mature fir, or that have a dense stocking of fir or a high proportion of fir in relation to other species. Vast budworm outbreaks in eastern North America, perhaps as many as 11 since 1704, have killed tens of millions of cubic meters (hundreds of millions of ft³) of balsam fir (6). Defoliation causes extensive root mortality. Evidence of budworm attack such as deformation, buried leaders, and decay can be seen 40 or more years later (1). Detailed articles about this important insect pest, with suggestions to alleviate damage, have been written (7,32) and a comprehensive bibliography assembled (25).
A classification system for tree vigor and budworm resistance was developed as a guide for selecting spruce and fir trees to remove or retain so as to make spruce-fir stands less vulnerable to spruce budworm attack. Silvicultural techniques designed to increase stand resistance to budworm cannot achieve their aim in the short term; several stand entries over the long term may be required, especially in stands dominated by balsam fir regeneration (46).
The balsam woolly adelgid (Adelges piceae), an introduced insect, is found in Southeastern Canada and in the Northeastern United States. Unless checked by low winter temperatures, populations build up and weaken or kill many trees. Severe stem attack can kill trees within 3 years. The insect also attacks twigs and buds, causing swellings and resulting in loss of new buds, gradual death of twigs and tops, and severe damage to regeneration. An abnormal growth of tracheids caused by insect saliva results in dark, brittle "redwood" (41).
The red heart fungus (Haematostereum sanguinolentum), causes much decay in living balsam fir. It enters almost entirely through injuries to the trunk and living branches (18). Losses from red heart rot are two or three times greater than those caused by butt rots (11,41). Six root and butt rots in balsam fir are economically important. These include the shoestring rot (Armillaria mellea), the two brown cubical rots (Tyromyces balsameus and Coniophora puteana), and the three white stringy rots (Poria subacida, Resinicium bicolor, and Scytinostroma galactinium). Another root disease of importance is Serpula himantioides. Phaeolus schweinitzii and Inonotus tomentosus also cause a small percentage of the root and butt rot in balsam fir (18). Mechanical or insect-caused wounds to the roots or basal areas of trees provide entrances for these fungi (41). Although the root and butt rots are not responsible for an excessive amount of cull in standing trees, they do weaken trees and make them more susceptible to wind damage, especially if trees are 20 cm (8 in) d.b.h. and larger. The defect caused by these rots is severe enough to be the decisive factor in setting the pathological rotation of fir at about 70 years (11,18,41).
Rot can begin in balsam fir as early as 40 years and increases as the trees get older. More than half generally are infected by the time they are 70 years old. No reliable external indicator of rot is known and even fruiting bodies are rare on living trees. Site seems to have an effect on the incidence and severity of rot; generally, the drier the site, the greater the damage from rot (41).
Specific causes of seedling diseases in nurseries have not been thoroughly reported. The foliage diseases of balsam fir are many but none are economically important to wood production. The same can be said for balsam fir's many stem or canker diseases (18).
The most conspicuous disease of balsam fir is witches' broom, caused by the rust fungus Melampsorella caryophyllacearum. Broomed shoots are upright and dwarfed and have yellow needles. Trunk and branch swellings are produced in the shoots (18).
Special Uses
The most important products made from balsam fir wood are pulpwood and lumber (43). The wood of balsam fir, as well as that of other true firs, is creamy white to pale brown. The sapwood has little odor or taste. Wood structure in the true firs is so similar that identification of species is impossible by examining only the wood (1,43).
Balsam fir is pulped by all of the pulping processes. Sulfate and semichemical processes are used most extensively. A fiber length of 3 to 4 mm A 12 to 0.16 in) is good, as is fiber quality. Because balsam fir is less dense than other major pulpwood species, its yield is lower (37).
The wood of balsam fir is light in weight, relatively soft, low in shock resistance, and has good splitting resistance. Recent testing of several mechanical properties of balsam fir and of red, white, and black spruce indicates strength values for balsam fir generally exceeding those of white spruce. In some tests, strength values were equivalent to or only slightly below the values of red and black spruce (5,34). Nail-holding capacity is low. Balsam fir is very low in resistance to decay (43). The major use of balsam fir lumber is for light-frame construction. Minor uses include paneling, crates, and other products not requiring high structural strength.
Balsam fir provides food or cover for some animals and both food and cover for others. Moose rely on balsam fir in winter when it is a major source of food. The use of balsam fir by deer for cover and shelter is well documented. During severe winter weather, especially in northern areas of the white-tailed deer range, lowland balsam fir stands and spruce-balsam fir swamps are used extensively as winter yarding areas. The fact that these sites usually contain, at best, only small amounts of preferred food suggests their attractiveness as shelter.
Other mammals use balsam fir to varying degrees. The snowshoe hare uses it for cover, and there is some seed and phloem feeding by various species of mice and voles. Red squirrels occasionally feed on balsam fir seed, bark, and wood. They prefer flower buds to vegetative buds. There is some use of wood by beaver for dam building, but little is used as food. Black bear strip bark and lick the exposed surfaces between bark and wood (1).
Balsam fir provides a minor part of the diet for both the spruce grouse and the ruffed grouse. Buds, tips, and needles are consumed, and more feeding occurs in winter than in summer. Thickets of balsam fir provide shelter for both birds (1). The response of bird populations to several forestry practices in stands containing balsam fir has been recorded (8,40). Species composition, the vertical and horizontal structure of the stand, and the extent of spruce budworm infestation influence the composition and density of bird populations.
Balsam fir is not widely planted as an ornamental nor does it offer much potential in areas other than northern New England, Canada, and perhaps the Lake States. Plantings as screens or as windbreaks are successful only when the moisture requirement of the species is met (1). On certain lands and especially on public lands, the unique presence of spruce-fir stands suggests management for esthetic values. In the southern Appalachian mountains, coniferous forests containing balsam fir are managed for watershed protection (44).
Oleoresin, a substance confined to the bark blisters of balsam fir, is used as a medium for mounting microscopic specimens and as a cement for various parts of optical systems. It is also used in the manufacture of medicinal compounds and spirit varnishes (4).
Balsam fir wood is not prized for fuelwood, but industries that use balsam fir for pulp and lumber products are using increasingly larger quantities of wood waste for the production of energy. The heating value of ovendry fir bark is 21 166 600 joules/kg (9,100 Btu/lb) (26).
The fir tree has been a favorite Christmas tree for more than 400 years. It remains among the top three species. In 1980, balsam fir ranked second behind Scotch pine (Pinus sylvestris), commanding 13.9 percent of the market (38). Sheared plantation-grown trees are usually preferred over wildings by retailers and consumers. Wreath-making is another holiday business that rivals that of Christmas tree sales in some areas. Prolonged needle retention after harvest, color, and pleasant fragrance are characteristics of balsam fir that make it attractive for these uses. Fragrance alone accounts for use of the needles as stuffing for souvenir pillows commonly sold in New England gift shops.
Genetics
Population Differences
Variation in balsam fir appears to be clinal and continuous and related to altitudinal gradient and to both east-west and north-south geographic gradients. Variation has been explored in a number of studies.
Balsam fir seedlings grown from seed collected along an elevational gradient in New Hampshire showed a clinal pattern of carbon dioxide uptake with respect to the elevational gradient. This suggests an adaption to temperature through natural selection (14). Another study failed to show that geographical variation in food quality of balsam fir needles is important to the spruce budworm diet but did suggest variation in food quality between locations (33).
In the southern Appalachians the monoterpenes- alpha-pinene and beta-phellandrene- appear to be the best taxonomic characteristics for separating balsam fir from Fraser fir, with alpha-terpene increasing southward and beta-terpene increasing northward. Because no regional variation pattern was evident for wood specific gravity or tracheid length, it has been suggested that only one species of balsam fir with three varieties be recognized in the Eastern United States: Abies balsamea var. balsamea, Abies balsamea var. phanerolepis, and Abies balsamea var. fraseri (29,39).
Balsam fir provenances from eastern portions of the range exhibited more vigor than those from western portions (24). This trait continued through 11 (22) and 13 years of total tree age (9). Southern sources tended to flush later, indicating selection for minimizing damage from the balsam gall midge (Dasineura balsamicola) and for resistance to late spring frost.
Specific gravity and tracheid length generally vary along an east-west gradient, with eastern sources of lower specific gravity and longer tracheids (9). Generally, trees from slow-growing sources have higher specific gravities and shorter tracheids than trees from fast-growing sources.
Races and Hybrids
No distinct races of balsam fir have been identified. Botanical varieties of balsam fir have been described, Abies balsamea var. phanerolepis being most important. This variety, the bracted balsam fir, is distinguished by its cone scales, which are shorter than the bracts. The variety phanerolepis is found infrequently from Labrador and Newfoundland to Maine and Ontario, and in the high mountains of New Hampshire, Vermont, and New York. It is found locally in northern Virginia and West Virginia (21,41,42), and commonly in several locations in Nova Scotia.
Until the late 1930's, natural or artificial hybrids of balsam fir had not been reported in North America. There were earlier reports, however, of hybrids between balsam fir and Siberian fir (Abies sibirica) in Europe (1).
Balsam fir is closely related to Fraser fir (A. fraseri). A taxon of doubtful status, A. intermedia, representing a possible cross between the two species, has been reported. This cross has also been reported as A. balsamea var. phanerolepis (1). Subalpine fir (A. lasiocarpa) also may hybridize with balsam fir where they adjoin in Alberta (42). Workers in Canada apparently have been successful in some instances in hybridizing balsam fir with several species of Abies, among them European silver fir (A. alba), alpine fir, and Fraser fir (1). Similar attempts in the United States have been only partially successful.
European horticulturists have propagated many forms of balsam fir for ornamental purposes. Plant form, needle color, and branch length and angle are characteristics usually manipulated. Nineteen such cultivars have been listed (1).
Literature Cited
- Bakuzis, E. V., and H. L. Hansen. 1965. Balsam fir-a monographic review. University of Minnesota Press, Minneapolis. 445 p.
- Barrett, J. W., ed. 1980. Regional silviculture of the United States. p. 25-65. John Wiley, New York.
- Baskerville, G. L. 1966. Dry-matter production in immature balsam fir stands: roots, lesser vegetation, and total stand. Forest Science 12(l):49-53.
- Bender, F. 1967. Canada balsam-its preparation and uses. Canada Department of Forestry and Rural Development Forestry Branch, Department Publication 1182. Ottawa, ON. 7 p.
- Bendtsen, B. Alan. 1974. Specific gravity and mechanical properties of black, red, and white spruce and balsam fir. USDA Forest Service, Research Paper FPL-237. Forest Products Laboratory. Madison, WI. 28 p.
- Blum, Barton M., David A. McLean. 1984. Chapter 6: Silviculture, forest management, and the spruce budworm. In Schmitt, Daniel M.; Grimble, David G.; Searcy, Janet L.; tech. coords: Managing the spruce budworm in eastern North America. Agric. Handb. 620. Washington, DC: U.S. Department of Agriculture: 83-10 1.
- Blum, Barton M., Harold M. Klaiber, and Arthur G. Randall. 1981. Northeastern spruce-fir. In Choices in silviculture for American forests. p. 10-15. Society of American Foresters, Washington, DC.
- Crawford, Hewlette S., and Richard W. Titterington. 1979. Effects of silvicultural practices on bird communities in upland spruce-fir stands. In Management of north central and northeastern forests for nongame birds. Workshop proceedings. p. 110-119. USDA Forest Service, General Technical Report NC-51. North Central Forest Experiment Station, St. Paul, MN.
- Dery, Patrick J., and Donald H. DeHayes. 1981. Variation in specific gravity and tracheid length among balsam fir provenances. In Proceedings, Twenty-seventh Northeastern Forest Tree Improvement Conference. p. 115-127. University of Vermont, Burlington. USDA Forest Service, Northeastern Forest Experiment Station, Broomall, PA.
- Eyre, F. H., ed. 1980. Forest cover types of the United States and Canada. Society of American Foresters, Washington, DC. 148 p.
- Frank, Robert M., and John C. Bjorkbom. 1973. A silvicultural guide for spruce-fir in the northeast. USDA Forest Service, General Technical Report NE-6. Northeastern Forest Experiment Station, Upper Darby, PA. 29 p.
- Frank, Robert M., and Barton M. Blum. 1978. The selection system of silviculture in spruce-fir stands-procedures, early results, and comparisons with unmanaged stands. USDA Forest Service, Research Paper NE-425. Northeastern Forest Experiment Station, Broomall, PA. 15 p.
- Frank, Robert M., and Lawrence 0. Safford. 1970. Lack of viable seeds in the forest floor after clearcutting. Journal of Forestry 68(12):776-778.
- Fryer, John H., and F. Thomas Ledig. 1972. Microevolution of the photosynthetic temperature optimum in relation to the elevational complex gradient. Canadian Journal of Botany 50:1231-1235.
- Ghent, A. W. 1958. Studies in forest stands devastated by the spruce budworm. Il. Forest Science 4:135-146.
- Griffin, Ralph H., and James E. Johnson. 1980. Polymorphic site index curves for spruce and balsam fir growing in even-aged stands in northern Maine. University of Maine Life Sciences and Agriculture Experiment Station, Bulletin 765. Orono. 22 p.
- Hampf, Frederick E. 1965. Site index curves for some forest species in the eastern United States (rev.). USDA Forest Service, Eastern Region, Upper Darby, PA. 43 p.
- Hepting, George H. 1971. Diseases of forest and shade trees of the United States. U.S. Department of Agriculture, Agriculture Handbook 386. Washington, DC. 658 p.
- Johnston, William F. 1986. Manager's handbook for balsam fir in the north central states. USDA Forest Service, General Technical Report NC-111. North Central Forest Experiment Station, St. Paul, MN. 27 p.
- Keays, J. L. 1975. Biomass of forest residuals. AlChE Journal 71(146):10-21.
- Lester, Donald T. 1968. Variation in cone morphology of balsam fir, Abies balsamea. Rhodora 80:83-94.
- Lester, D. T., C. A. Mohn, and J. W. Wright. 1976. Geographic variation in balsam fir: 11-year results in the Lake States. Canadian Journal of Forest Research 6:389-394.
- Little, Elbert L., Jr. 1979. Checklist of United States trees (native and naturalized). U.S. Department of Agriculture, Agriculture Handbook 541. Washington, DC. 375 p.
- Lowe, W. J., H. W. Hocker, Jr., and M. L. McCormack, Jr. 1977. Variation in balsam fir provenances planted in New England. Canadian Journal of Forest Research 7:63-67.
- McKnight, M. E., D. T. Jennings, S. C. Hacker, and F. B. Knight. 1988. North American coniferaphagous choristoneura: a bibliography. USDA Forest Service, Bibliographies and Literature of Agric. No. 59. Washington, DC. 580 p.
- Millikin, D. E. 1955. Determination of bark volumes and fuel properties. Pulp and Paper Magazine of Canada 56(13):106108.
- Page, G. 1976. Quantitative evaluation of site potential for spruce and fir in Newfoundland. Forest Science 22:131-143.
- Randall, Arthur G. 1976. Natural regeneration in two spruce-fir clearcuts in eastern Maine. University of Maine, Research in the Life Sciences 23(13). Orono. 10 p.
- Robinson, John F., and Eyvind Thor. 1969. Natural variation in Abies of the southern Appalachians. Forest Science 15:238-245.
- Rudolf, Paul 0. 1966. Botanical and commercial range of balsam fir in the Lake States. USDA Forest Service, Research Note NC-16. North Central Forest Experiment Station, St. Paul, MN. 4 p.
- Safford, Lawrence 0. 1968. Ten-year average growth rates in the spruce-fir region of northern New England. USDA Forest Service, Research Paper NE-93. Northeastern Forest Experiment Station, Upper Darby, PA. 20 p.
- Sanders, C. J.; Stark, R. W.; Mullins, E. J.; Murphy, J., eds. 1985. Recent advances in spruce budworm, research. Proceedings, CANUSA spruce budworms research symposium, Bangor, ME, Sept. 16-20. Ottawa, ON: Canadian Forestry Service: 527 p.
- Shaw, G. G., and C. H. A. Little. 1977. Natural variation in balsam fir foliar components of dietary importance to spruce budworm. Canadian Journal of Forest Research 7:47-53.
- Sinclair, Steve, Bob Garett, and Jim Bowyer. 1981. Potential lumber grade yields from balsam fir and spruce budworm killed balsam fir. Timber Producers Bulletin 36: 10-11.
- Solomon, Dale S. and Robert M. Frank. 1983. Growth response of managed uneven-aged northern conifer stands. USDA Forest Service, Research Paper NE-517. Northeastern Forest Experiment Station, Broomall, PA. 17 p.
- Solomon, Dale S., Richard A. Hosmer, and Homer T. Hayslett, Jr. 1987. FIBER handbook: a growth model for spruce-fir and northern hardwood types. USDA Forest Service, Research Paper NE-602. Northeastern Forest Experiment Station, Broomall, PA. 17 p.
- Sonderman, David L. 1970. Balsam fir. USDA Forest Service, American Woods Series FS-234. Washington, DC. 8 p.
- Svinicki, Jane A. 1981. Five-year over-view of retail Christmas tree market. p. 6-8. 1981 Christmas Merchandiser, National Christmas Tree Association, Milwaukee, WI.
- Thor, Eyvind, and Paul E. Barnett. 1974. Taxonomy of Abies in the southern Appalachians: variations in balsam monoterpenes and wood properties. Forest Science 20:32-40.
- Titterington, R. W., H. S. Crawford, and B. N. Burgason. 1979. Songbird responses to commercial clearcutting in Maine spruce-fir forests. Journal of Wildlife Management 43(3):602-609.
- USDA, Forest Service. 1965. Silvics of forest trees of the United States. H. A. Fowells, comp. U.S. Department of Agriculture, Agriculture Handbook 271. Washington, DC. 762 p.
- USDA, Forest Service. 1974. Seeds of woody plants in the United States. C. S. Schopmeyer, tech. coord. U.S. Department of Agriculture, Agriculture Handbook 450. Washington, DC. 883 p.
- USDA, Forest Service. 1974. Wood handbook: wood as an engineering material. U.S. Department of Agriculture, Agriculture Handbook 72 (rev.). Washington, DC. 439 p.
- Walker, Lawrence C. 1967. Silviculture of the minor southern conifers. p. 56-62. In Stephen F. Austin State College School of Forestry, Bulletin 15. Nacogdoches, TX.
- Weetman, G. F., W. W. Grapes, and G. J. Frisque. 1973. Reproduction and ground conditions 5 years after pulpwood harvesting: results from 37 study areas in eastern Canada. Pulp and Paper Research Institute of Canada LRR/51. Pointe Claire, PQ. 97 p.
- Westveld, Marinus. 1954. A budworm vigor-resistance classification for spruce and balsam fir. Journal of Forestry 52:11-24.